La Enfermedad de las Postlarvas Translúcidas más allá de China: desde la identificación del brote hasta la preparación para el diagnóstico
Estudios recientes están proporcionando a los laboratorios de producción de postlarvas de camarón herramientas más sólidas para reconocer, caracterizar y detectar la enfermedad de postlarvas translúcidas (TPD) en el camarón blanco del Pacífico
Cuando los laboratorios de producción de postlarvas sufren pérdidas, estas pueden escalar rápidamente. Un tanque que parece estar bajo control por la mañana puede deteriorarse drásticamente al final del día, dejando al personal con poco tiempo para reaccionar y aún menos para determinar la causa o causas del problema. Esto es lo que convierte a la enfermedad de postlarvas translúcidas (TPD) en una preocupación tan grave. Afecta a los camarones en una de las etapas de producción más vulnerables, cuando los animales son frágiles, las decisiones sobre su traslado requieren rapidez y un diagnóstico tardío puede traducirse directamente en pérdidas económicas significativas.
Desde su reporte inicial en China en 2020, la TPD ha pasado de ser una enfermedad emergente a representar un desafío importante para los laboratorios de producción, con repercusiones relevantes para la producción, el traslado de animales y la bioseguridad. Tres estudios recientes de nuestro grupo contribuyen a llevar el debate más allá del simple reconocimiento de brotes hacia un marco más útil, basado en la causalidad, la patología y una detección fiable. En conjunto, estos estudios abordan tres cuestiones fundamentales para las operaciones de los laboratorios: ¿qué causa estas mortalidades?, ¿qué tipo de daño tisular provoca el patógeno? y ¿pueden los laboratorios analizar las poblaciones antes de trasladar los animales a otro laboratorio, centro de pre-cría o granja?
Este artículo es una adaptación y resumen de tres publicaciones originales (PLOS One; Microbial Genomics; Journal of Microbiological Methods) que señalan un problema concreto relacionado con Vibrio parahaemolyticus, un sistema de virulencia específico asociado a toxinas y un enfoque de vigilancia basado en una técnica validada de reacción en cadena de la polimerasa cuantitativa (qPCR, una técnica que combina la amplificación de una secuencia de ADN diana con la cuantificación de la concentración de dicha especie de ADN en la reacción). En términos prácticos, esto significa que el TPD ahora puede entenderse no solo como un síndrome observado en campo, sino también como una enfermedad con una base biológica más clara y un conjunto creciente de herramientas para la toma de decisiones en los centros de producción de semilla.
Diseño del estudio
El primer estudio comenzó con un caso clínico proveniente de un criadero de camarones en el Sudeste Asiático, fuera de China. El criadero experimentaba una mortalidad repentina y grave en postlarvas de camarón blanco del Pacífico (Penaeus vannamei), con pérdidas acumuladas superiores al 70 por ciento en un plazo de cinco días. Los animales afectados presentaban los signos actualmente asociados a la TPD: hepatopáncreas pálido, tracto digestivo pálido o vacío y cuerpo translúcido.
![Ad for [Aquademia]](https://www.globalseafood.org/wp-content/uploads/2025/07/aquademia_web2025_1050x125.gif)
A partir de estas postlarvas enfermas, aislamos una cepa bacteriana denominada AG1 y la identificamos como V. parahaemolyticus. Este hallazgo fue importante porque constituyó el primer informe confirmado de una cepa de este Vibrio asociada a la TPD (VTPD) en camarones procedentes de fuera de China. Esto amplía significativamente el alcance de la información sobre la TPD. La enfermedad ya no es relevante únicamente en el contexto de los informes iniciales, sino también para los criaderos fuera de China que podrían enfrentarse a riesgos de bioseguridad similares.
El segundo estudio analizó la misma cepa a nivel genómico. La secuenciación del genoma completo reveló que la cepa AG1 posee un genoma de aproximadamente 5,5 Mb, organizado en dos cromosomas y tres plásmidos. Uno de estos plásmidos portaba tres genes asociados a la virulencia – vhvp-1, vhvp-2 y vhvp-3 – los cuales han sido vinculados a la TPD. Este estudio se diseñó para esclarecer el mecanismo patogénico de la cepa y compararla con las cepas de TPD descritas anteriormente en China.
El tercer estudio se centró en una necesidad práctica compartida por criaderos y laboratorios de diagnóstico: la detección fiable. En este trabajo, se desarrollaron tres ensayos de PCR en tiempo real tipo TaqMan (sondas de hidrólisis diseñadas para aumentar la especificidad de la PCR cuantitativa) dirigidos a los genes vhvp-1, vhvp-2 y vhvp-3, siguiendo el marco de desarrollo y validación de ensayos establecido por la Organización Mundial de Sanidad Animal (OMSA/WOAH). El objetivo era proporcionar una herramienta más fiable para el diagnóstico, el cribado rutinario y la bioseguridad relacionada con el movimiento de animales.
Resultados y discusión
Una de las conclusiones más claras del primer estudio fue que la cepa AG1 se ajusta al perfil de la TPD (enfermedad de necrosis aguda del hepatopáncreas transmitida por *Vibrio*) más que al de la enfermedad de necrosis aguda del hepatopáncreas (AHPND). El cribado mediante PCR detectó vhvp-1 y vhvp-2, pero el aislado carecía de los genes de las toxinas pirA/pirB asociados a la AHPND. Esta distinción es importante porque los episodios de mortalidad temprana en los criaderos a menudo se clasifican bajo términos generales como “vibriosis” o se asume que siguen patrones de diagnóstico más conocidos. Nuestros hallazgos demuestran que la TPD no debe considerarse simplemente otra forma de AHPND. Aunque ambas están asociadas a Vibrio, implican sistemas de virulencia diferentes y, por tanto, requieren dianas diagnósticas distintas.
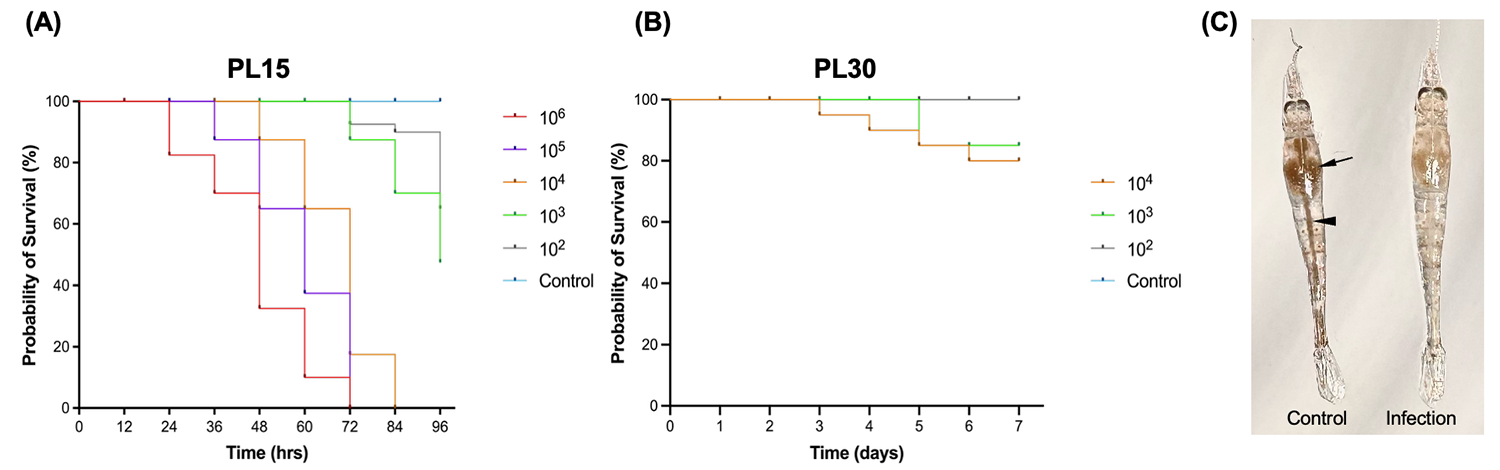
Los ensayos de infección confirmaron además la elevada virulencia de la cepa AG1. En camarones PL15, la exposición por inmersión provocó una mortalidad marcada y dependiente de la dosis, con una CL50 de 8,51 × 10² UFC/mL a las 96 horas. Por el contrario, los camarones PL30 de mayor tamaño fueron menos susceptibles, lo que sugiere que el tamaño del huésped o su etapa de desarrollo influyen en la evolución de la enfermedad. Para los centros de producción de semilla (hatcheries), este es uno de los hallazgos prácticos más útiles. Las etapas de poslarva más tempranas parecen ser las más vulnerables, por lo que constituyen el periodo de mayor prioridad para el monitoreo y el cribado.
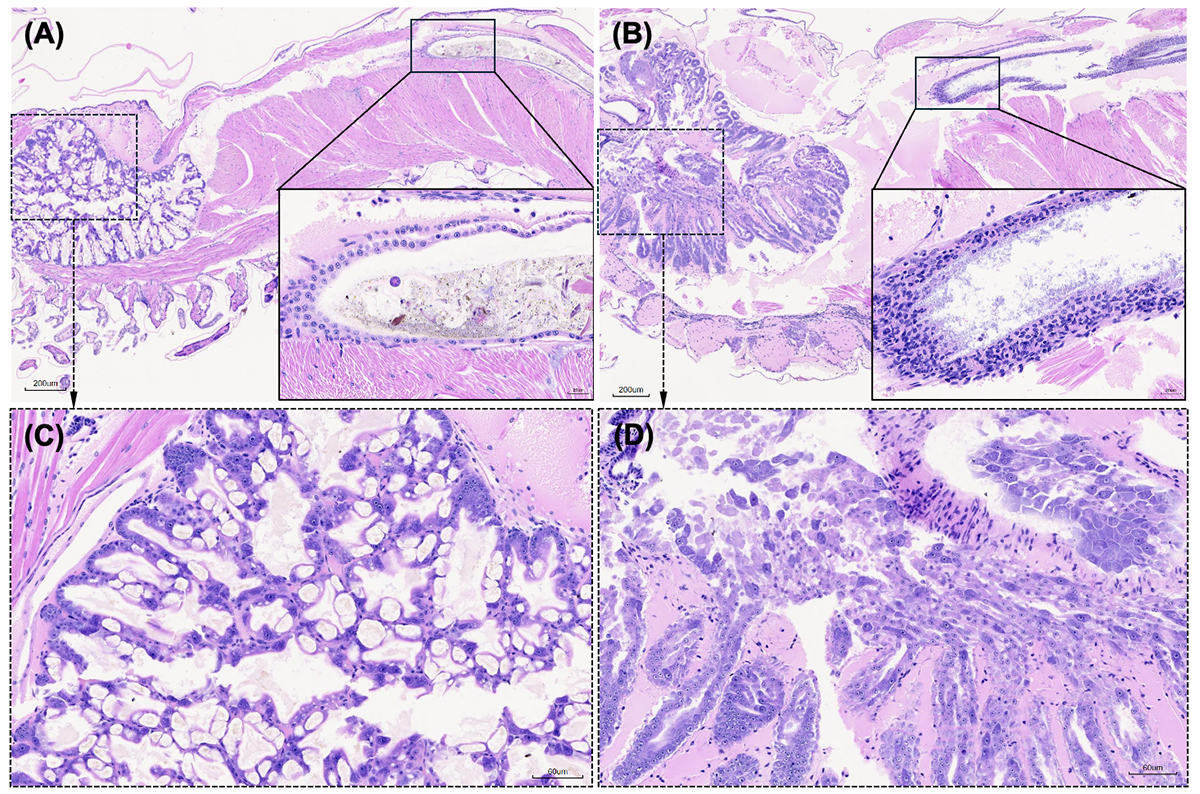
La histopatología añadió otra dimensión importante al perfil de la enfermedad. Las postlarvas infectadas mostraron degeneración hepatopancreática caracterizada por necrosis tubular, desprendimiento de células epiteliales de los túbulos e invasión bacteriana. Sin embargo, las lesiones no se limitaron al hepatopáncreas. Un hallazgo adicional notable fue la enteritis hemocítica, caracterizada por la pérdida del epitelio de la mucosa intestinal, una inflamación marcada y la formación de una capa gruesa de hemocitos en el intestino. Estas observaciones sugieren que la TPD no es simplemente una enfermedad hepatopancreática. El intestino también parece ser un órgano diana importante, y esta afectación tisular más amplia podría ayudar a explicar la rapidez y gravedad con las que progresa la enfermedad en los camarones poslarva tempranos, que son más frágiles.
El estudio del genoma proporcionó una segunda línea de evidencia de que este patógeno es biológicamente distinto. La cepa AG1 portaba sus tres genes asociados a la TPD en un plásmido de aproximadamente 69,7 kb. Aunque este plásmido es más pequeño que el plásmido de TPD de mayor tamaño descrito anteriormente en China, conserva la región asociada a la virulencia. Los genes vhvp en AG1 mostraron una gran similitud con los de la cepa de referencia china, y las proteínas codificadas contenían dominios consistentes con un complejo de toxinas tipo Tc. En términos prácticos, esto indica que la TPD está asociada a un sistema de toxinas especializado, en lugar de a las toxinas pirA/B (más conocidas) implicadas en la AHPND. Esta distinción refuerza la base biológica para tratar la TPD como una entidad patológica independiente y subraya la importancia de centrarse en los marcadores de virulencia correctos en los ensayos de diagnóstico.
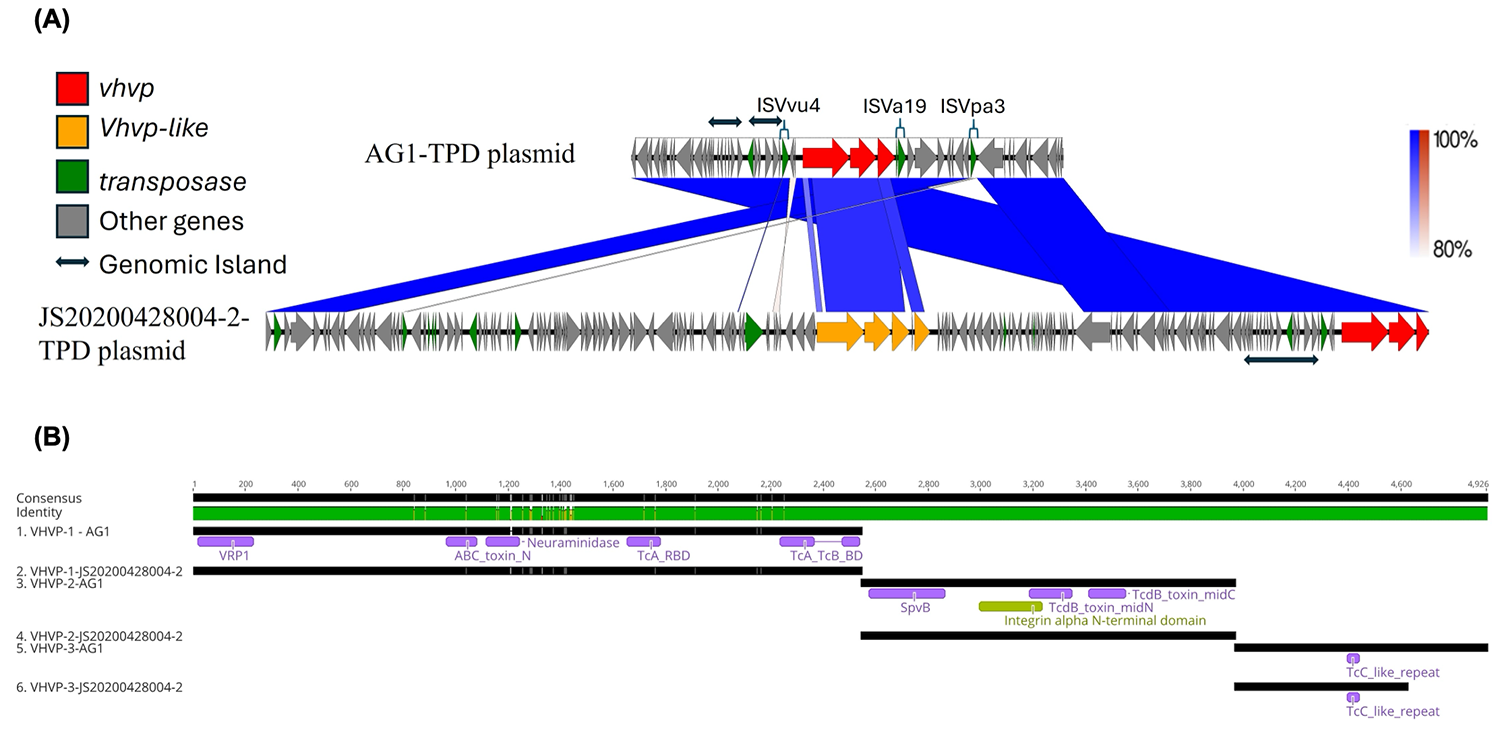
Los datos genómicos también sugieren que las cepas asociadas a la TPD no son genéticamente estáticas. Aunque la cepa AG1 estaba estrechamente relacionada con cepas chinas descritas anteriormente, no era idéntica a ellas. Las diferencias en la estructura del plásmido, las islas genómicas y las características relacionadas con la transferencia indican que estas bacterias pueden seguir evolucionando y, al mismo tiempo, conservar el conjunto de genes de virulencia asociados a la patogenicidad. Para la industria camaronera, esto constituye un recordatorio importante de que las enfermedades bacterianas emergentes son dinámicas. Las estrategias de vigilancia deben evolucionar a la par de los patógenos que están diseñadas para detectar.
El tercer estudio tradujo estos hallazgos biológicos en una herramienta de diagnóstico práctica. Los tres ensayos TaqMan recién desarrollados mostraron una sensibilidad y una especificidad diagnósticas del 100 por ciento, con un límite de detección de 10 copias por reacción. Asimismo, los ensayos no presentaron reactividad cruzada con camarones libres de patógenos específicos (SPF) ni con camarones infectados por otros patógenos importantes, incluidos EHP, NHP, IHHNV, WSSV y VpAHPND. Para los centros de producción de semilla (hatcheries) y los laboratorios de diagnóstico, este nivel de rendimiento es de gran relevancia. Un ensayo de cribado solo resulta valioso si es capaz de detectar de forma fiable el objetivo previsto sin confundir la TPD con otros patógenos comunes en la producción de camarón.
Desde la perspectiva de la validación, los ensayos también demostraron una sólida linealidad, una eficiencia aceptable y una precisión coherente con el uso diagnóstico rutinario. Aunque se trata de parámetros técnicos, su implicación práctica es clara: los ensayos están superando la fase de prueba de concepto para adquirir valor operativo como herramientas de cribado en centros de producción de semilla, programas de cría y laboratorios de sanidad de animales acuáticos. Su mayor valor podría residir en la etapa previa al traslado de las postlarvas, momento en el que prevenir la propagación de patógenos resulta mucho más eficaz que intentar controlarlos una vez introducidos en un nuevo sistema.
Dinh-Hung, TPD, Tabla 1
| Cebadores/sondas | Conjuntos | Target | Sequencia (5'-3') | |
|---|---|---|---|---|
| V1-qF | V1 | vhvp-1 | AATGTAGCCTCCATATCGGAAATC | |
| V1-probe | – | – | 6FAM-CATCAATTGCTTGCACCTCACATCGC-TAMRA | |
| V2-qF | V2 | vhvp-2 | AATCGCCACTCACTCAGATTCA | |
| V2-qR | – | – | ACGCCCCGCTTTTTTGTT | |
| V2-probe | – | – | 6FAM-CAGCTCAGCGTTATCCAGTCGCGA-TAMRA | |
| V3-qF | V3 | vhvp-3 | ACTACCGCCACCCTGATGAG | |
| V3-qR | – | – | GCGCCACGTGTATCGTACTG | |
| V3-probe | – | – | 6FAM-CAAATACGACACAGACACGCATTACGCATC-TAMRA |
Para los gerentes de laboratorios de producción de semilla (hatcheries) y los productores, destacan tres mensajes prácticos. En primer lugar, la TPD debe considerarse una amenaza real para la bioseguridad en las etapas tempranas de vida. Cuando las postlarvas presentan hepatopáncreas pálido, intestino vacío y cuerpo translúcido, y cuando la mortalidad aumenta rápidamente, se debe considerar inmediatamente la TPD en la investigación diagnóstica. En segundo lugar, no todos los brotes asociados a Vibrio son biológicamente equivalentes. Las denominaciones generales como “vibriosis” pueden ser útiles en campo, pero pueden ocultar diferencias importantes que afectan al diagnóstico y al control. En tercer lugar, el cribado previo al traslado puede ser una de las aplicaciones más valiosas de los nuevos ensayos de qPCR. Estas herramientas permiten a los laboratorios utilizar los análisis no solo para investigar la mortalidad una vez que esta ha comenzado, sino también para reducir el riesgo de transferir poblaciones infectadas a nuevos sistemas de producción.
Perspectivas
En conjunto, los tres estudios aquí resumidos ofrecen una visión mucho más precisa de la TPD. Uno de ellos confirma que una cepa de V. parahaemolyticus asociada a la TPD está presente fuera de China y es capaz de causar una mortalidad severa en postlarvas de P. vannamei. Un segundo estudio demuestra que esta cepa posee una arquitectura genómica distintiva asociada a toxinas, vinculada a vhvp-1, vhvp-2 y vhvp-3. Un tercer estudio aporta ensayos TaqMan validados que pueden respaldar el diagnóstico, la vigilancia y el cribado previo a traslados.
Para la industria camaronera, estos hallazgos cambian la naturaleza del debate. La TPD ya no es solo la descripción de un brote; ahora es una enfermedad con una base causal más clara, un perfil patológico y genómico mejor definido y una estrategia de detección práctica. Estos avances no eliminan el riesgo, pero proporcionan a los laboratorios una base más sólida para una detección temprana, análisis más fiables y medidas de bioseguridad más estrictas orientadas a prevenir mayores pérdidas.
Ahora que Ud. ha llegado al final del artículo...
… por favor considere apoyar la misión de la GSA de promover prácticas responsables de productos del mar a través de la educación, la defensa y las garantías de terceros. El Advocate tiene como objetivo documentar la evolución de las prácticas responsables de productos del mar y compartir el amplio conocimiento de nuestra amplia red de colaboradores.
Al hacerse miembro de la Global Seafood Alliance, está asegurando de que todo el trabajo pre-competitivo que hacemos a través de los beneficios, recursos y eventos para miembros pueden continuar. La membresía individual cuesta solo $50 al año.
¿No es miembro de la GSA? Únase a nosotros.
Authors
-

Aquaculture Pathology Laboratory
The University of Arizona
Tucson, AZ 85721 USA -

Aquaculture Pathology Laboratory
The University of Arizona
Tucson, AZ 85721 USA -

Corresponding author
Aquaculture Pathology Laboratory
The University of Arizona
Tucson, AZ 85721 USA
Related Posts

Health & Welfare
Calidad, supervivencia de la progenie de L. vannamei de hembras sometidas a ablación y a no-ablación
Evaluación de la calidad y supervivencia de postlarvas de camarón blanco del Pacífico de reproductores hembras no sometidas a ablación, una estrategia holística de bioseguridad y gestión.

Health & Welfare
AHPND es una enfermedad crónica del camarón blanco del Pacífico de América Latina
Hay una nueva fase de infección para la enfermedad de Necrosis Hepatopancreática Aguda en las granjas camaroneras de América Latina, en contraste con las observaciones del sudeste asiático.

Health & Welfare
Cuatro cepas de AHPND identificadas en granjas de camarón de América Latina
Se conocen dos genes de virulencia que codifican una toxina binaria de Photorhabdus relacionada con insectos que causa la enfermedad de necrosis hepatopancreática aguda en camarones. Las patogenicidades de estas cepas de V. campbellii fueron evaluadas mediante infección de laboratorio y posterior examen histológico en camarones P. vannamei.

Health & Welfare
Probióticos benefician al camarón blanco del Pacífico desafiado con AHPND
Un estudio fue llevado a cabo para medir los efectos de probióticos comerciales en camarón blanco del Pacífico en un modelo de desafío a AHPND estandarizado bajo condiciones controladas de laboratorio. Los resultados muestran que los tratamientos de probióticos por sí mismos tienen efectos beneficiosos, como una mayor supervivencia y signos histológicos de regeneración de hepatopáncreas.