Effects of dietary lipid on the gut microbiota of first feeding Atlantic salmon
Fish mean weights, total lengths were not significantly different between diets based on dietary fish oil, vegetable oil

A variety of alternative feed ingredients is being increasingly used in aquafeeds, and because changes in diet ingredients can alter the gut microbiota of farmed aquatic species, it is important to evaluate the impact of these new diets – with lower fishmeal and fish oil contents – on the composition of the gut microbial communities for aquacultured species.
Although feed is considered the main factor affecting the gut bacterial communities in fish species, data from previous studies have also shown variations in gut microbial communities across development stages, which seem to be affected not only by the provided feed but also from the microbial communities of the rearing environment. For example, recent research reported a strong association between the environment (i.e., tank biofilm and water from the hatchery installation) and Atlantic salmon (Salmo salar) mucosal microbiota. And in rainbow trout, gut microbiota was detectable before first feeding commenced, potentially due to contact with the surrounding water and yok sac digestion, indicating that gut microbiota establishment initiates at first feeding and that diet type affects the bacterial composition.
Other than a few studies investigating the gut microbial communities in early life stages of Atlantic salmon, there is a knowledge gap on the effects of a different dietary lipid source on the gut microbiota in early life stages of this fish species.
This article – adapted and summarized from the original publication [Nikouli, E. et al. 2021. Dietary Lipid Effects on Gut Microbiota of First Feeding Atlantic Salmon (Salmo salar L.). Front. Mar. Sci., 20 May 2021] – evaluated the influence of total replacement of dietary fish oil with a blend of terrestrial alternative oils (rapeseed, linseed and palm oils) on the intestinal microbiota of first feeding Atlantic salmon. This research also characterized the bacterial communities of the rearing environment to determine their contribution in the early colonization and the succession of the fish intestines.
![Ad for [BSP]](https://www.globalseafood.org/wp-content/uploads/2025/07/BSP_B2B_2025_1050x125.jpg)
Study setup
The study was conducted at the Ervik Hatchery (Frøya, Norway), where a fast-growing Atlantic salmon aquaculture strain was reared from fertilized eggs until 93 days post first feeding (dpff). The two diets used in this study, FD (fish oil diet) and VD (blend of rapeseed, linseed and palm oils diet), were formulated to cover the fish nutritional requirements. Other than the lipid sources, the rest of the feed components were identical in the two diets.
Each dietary treatment was tested in duplicated groups of 200 Atlantic salmon individuals (0.23 grams ± 0.03 grams per fish). Fish hindgut samples were collected at 0, 35, 65 and 93 dpff. Samples of fertilized eggs, yolk sac larvae, rearing water and feed were also collected to assess a possible contribution of their microbiota to the colonization and bacterial succession of the fish intestines.
For detailed information on experimental design and sampling; DNA extraction and sequencing; and data analyses, refer to the original publication.
Results and discussion
We evaluated the influence of dietary administration of two different lipid sources (fish oil and vegetable oil) on the gut bacterial communities of first feeding Atlantic salmon. Moreover, we characterized the bacterial communities from the rearing environment (rearing water and feeds) and the epibiotas of fertilized eggs and yolk sac larvae to determine their contribution in the bacterial colonization and succession of the gut. Previous studies suggest that the bacterial communities of the rearing environment, mainly from the rearing water and the feed, are important sources for community assembly of the intestinal microbiota of fish. However, our results suggest that substitution of fish oil by vegetable oils did not significantly affect the composition of intestinal microbial communities in the same host species.
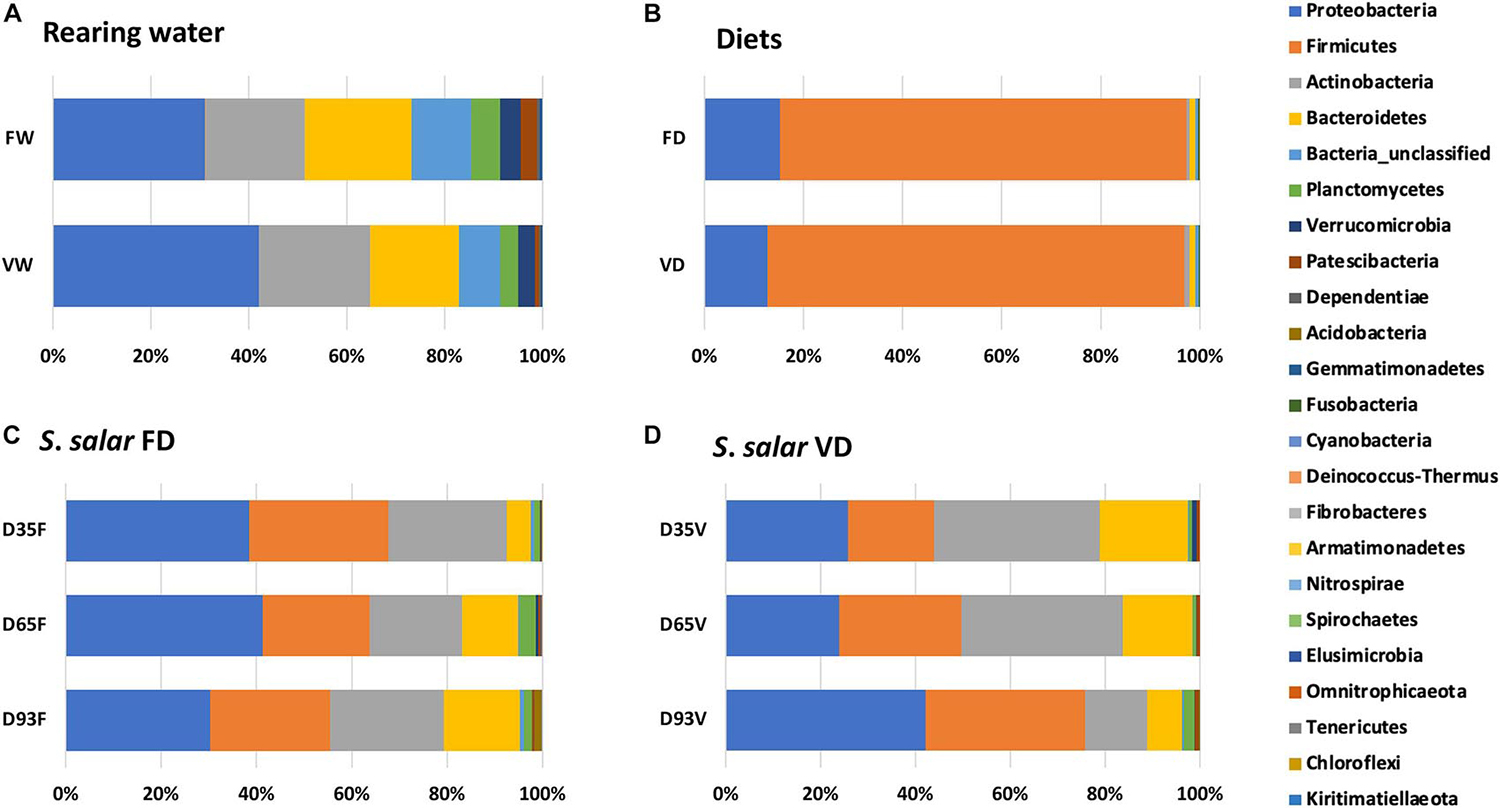
Furthermore, our results indicate little overlap between the bacterial communities of the host with that of the rearing environment (water and feed), whereas the life stage appeared to be the main factor affecting the structure of gut microbiota. These results agree with previous findings from other researchers, who studied 96 wild-caught Atlantic salmon individuals with different age and habitats and reported groupings of their intestinal bacterial communities based on the lifecycle stage. Another group of researchers reported stage-specific microbial enrichment in intestinal mucosa of the same host species (samples from embryonic stages up to 80 weeks post-hatch).
Similar stage-specific signatures have also been reported across development in other fish species, further supporting that the life stage seems to be the primary force shaping gut microbiota in juvenile stages of fish. The change in microbiota with life stage can be due to both host-microbe (e.g., development in morphology and immune system) and microbe-microbe interactions (mutualism, commensalism and competition). The significance of these factors is, however, still not known.
Proteobacteria, Firmicutes, Actinobacteria and Bacteroidetes were the dominant bacterial phyla detected in the host samples for both dietary treatments in our study. These bacterial phyla seem to characterize the bacterial communities in individuals of Atlantic salmon at the freshwater life cycle stages. These bacterial phyla are also commonly found in the gut bacterial communities of both saltwater and freshwater fish species.
Despite the fact that the two experimental feeds contained almost entirely Firmicutes, the increase in relative abundance of Firmicutes in samples after the onset of feeding was not solely due to feed-specific operational taxonomic units, OTUs [used to classify groups of closely related individuals]. It should also be noted that 26.4 percent of the bacterial representatives detected on fertilized eggs (EG) were not detected in the water of the incubation tank (WST). This supports the view that the microbial communities of fish eggs may be vertically transmitted from their parents or horizontally from their breading tank.
We also found that, in agreement with previous studies by various researchers, the observed species richness in water samples was always an order of magnitude higher than the richness of the host samples. Bacterial communities in rearing water did not show major shifts during our experiment.
![Fig. 2: Venn diagram [used to show similarities and differences between different groups] demonstrating the number of the shared and unique operational taxonomic units (OTU) and their percentage of the total library, between Atlantic salmon samples [pre-feeding (EG,YS, D0) and post first feeding stages (D35, D65, D93 in both diets)], diets (FD and VD) and rearing water sample (STW, FW, VW). Adapted from original.](https://www.globalseafood.org/wp-content/uploads/2021/09/NIKOULI-Fig-2-300x245.jpg)
In general, our findings indicate that when feeding on either fish oil or vegetable oil-based diets, the Atlantic salmon growth variables of mean wet weight and total length did not differ significantly during our experiment (p > 0.05). No significant differences were found between the two dietary groups, regarding their gut bacteria composition, after analysis of their sequencing data. Instead, gut microbiota changed with age, and each stage was characterized by different dominant bacteria. These operational taxonomic units (OTUs) were related to species that provide different functions and have been isolated from a variety of environments. The results also show little OTUs overlap between the host and rearing environment microbiota. Overall, the results of our study revealed the occurrence of a core microbiota in early life of Atlantic salmon independent of the origin of the oil used in their feeds.
Perspectives
Our study evaluated the effect of total fish oil replacement by a blend of terrestrial vegetable oils (rapeseed, linseed and palm oils) in the feed on the colonization and the bacterial succession in first feeding of Atlantic salmon, up to 93 days post first feeding (dpff).
We demonstrated that feeding on either fish oil or terrestrial vegetable oil diets did not result in significant differences in the intestinal gut microbiota and growth performance parameters (wet weight and total length). On the contrary, the composition of gut microbiota changed with age, and each stage was characterized by different dominant bacteria. These OTUs are related to species that may have probiotic activity on the host. Our study also revealed the occurrence of a core microbiota independent of the studied life stages and diet of the fish.
Our results indicate that total fish oil replacement in Atlantic salmon with terrestrial vegetable oils is feasible and can lead to lower-cost formulated feeds. Future work should aim to understand the functional role of the detected core bacterial community, which could lead to further growth performance and host-health optimization.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Corresponding author
Department of Ichthyology and Aquatic Environment, School of Agricultural Sciences, University of Thessaly, Volos, Greece; and Department of Biotechnology and Food Science, Norwegian University of Science and Technology (NTNU), Trondheim, Norway -
Department of Ichthyology and Aquatic Environment, School of Agricultural Sciences, University of Thessaly, Volos, Greece
-
Department of Biology, Norwegian University of Science and Technology (NTNU), Trondheim, Norway
-
Department of Biology, Norwegian University of Science and Technology (NTNU), Trondheim, Norway
-
Department of Biotechnology and Food Science, Norwegian University of Science and Technology (NTNU), Trondheim, Norway
-
Department of Biotechnology and Food Science, Norwegian University of Science and Technology (NTNU), Trondheim, Norway