Investigating the potential for domestication selection in the Eastern oyster
Study tests starvation resistance of larvae from wild and artificially selected broodstock

One concern with selective breeding as part of a population management strategy is that captivity itself can impose unintended artificial selection. Evolutionary responses to this “domestication selection” can be swift in captive populations with the potential for reduced fitness in the wild relative to wild-born individuals.
When domestication selection is strong within a single propagation cohort, such as was demonstrated for salmonids, then its effects can potentially impact the success of hatchery-based population supplementation. These considerations are of particular relevance to marine bivalves because (1) they express extreme versions of this life history, (2) native bivalve populations are depleted in some areas and receiving hatchery-based population supplementation and (3) selectively bred lines intended for commercial aquaculture have been promoted and used for population supplementation, for example in an attempt to mitigate disease mortality.
In the United States, Eastern oyster (Crassostrea virginica) restoration approaches often include the hatchery production and planting of spat (juvenile oysters) on shell to supplement wild recruitment and help jumpstart a population on restored habitat. Oysters produced for the aquaculture industry are often selectively bred for traits that speed up production, such as fast growth, and that improve survival (e.g. disease resistance). However, it is unclear whether or not other (unintended) traits are evolving due to genetic correlations, adaptation to the artificial environment, or heritable epigenetic changes during hatchery culture.
This article – adapted and summarized from the original publication – reports on a study to examine the potential effects of domestication selection in oysters. The experiment compared the starvation resistance of oyster larvae produced from wild (no prior hatchery exposure) and artificially selected (over multiple generations of hatchery propagation) broodstock oysters.
Study setup
To test for domestication selection in marine bivalves, we focused on a fitness-related trait (larval starvation resistance) that could be altered under artificial selection. Using larvae produced from a wild population of C. virginica and a selectively bred, disease-resistant line we measured growth and survival during starvation versus standard algal diet conditions.
Wild adult oysters were collected from the Choptank River, Maryland in the Chesapeake Bay and two disease-resistant aquaculture lines (Deby (DBY) and DBY-CROSbreed (XB)) were obtained from the Virginia Institute of Marine Science, Aquaculture Genetics and Breeding Technology Center (ABC). These lines have been bred over multiple generations with hatchery propagation and intensive selection for disease resistance.
Broodstock were conditioned and spawned and the oysters were allowed to finish spawning in their individual container to collect and isolate gametes for each individual. A total of six pair-cross fertilizations were completed between two females and three males for each strain type (wild and aquaculture) within one hour of the start of spawning to assure quality of gametes. The DBY and XB selection lines are maintained with methods that limit inbreeding, but to eliminate any potential for inbreeding effects here, our experimental aquaculture cohort was created with males from the DBY line and females from the XB line (hereafter referred to as AQF1).
The oyster larvae were reared in 200-liter tanks at a density of 30 larvae per mL for seven days and were fed a diet of 50:50 Isochrysis galbana and Chaetoceros calcitrans beginning at 10,000 cells per mL on day one and increased each day by 10,000 cells per mL. Water changes were completed on day three and then every other day thereafter with no culling until day seven. Then, an experiment was conducted to measure growth and survival during starvation versus standard algal diet conditions.
For detailed information on the experimental design; broodstock conditioning and spawning; total organic carbon (TOC) analysis; starvation challenge; respiration rate measurements; and statistical analyses, refer to the original publication.
Results and discussion
Based on our results and those of other researchers, we hypothesize that the ability to absorb, transport and assimilate micronutrients may be an important source of differentiation between the lines tested and represent promising phenotypes to explore as possible mechanistic changes associated with domestication selection.
It is plausible that aquaculture lines can adapt to the hatchery environment, in which food is provided in excess, thereby limiting their ability to utilize dissolved organic matter (DOM) during prolonged starvation events as a result of many generations of hatchery propagation. However, this requires further and more detailed testing to understand the role of DOM under food limitation.
Regarding survival during prolonged starvation, the most dramatic mortality was observed between days 8 and 10 of the starvation (age 15 to 17 days) in the AQF1 line suggesting that a critical physiological threshold was reached. This is consistent with the continued mortality we observed during the recovery period, but clearly there was individual variation in this threshold because some larvae did recover. Survival continued to decline through age 22 days (five days into the recovery), during which growth also remained stunted, suggesting that larvae had not yet rebounded physiologically.
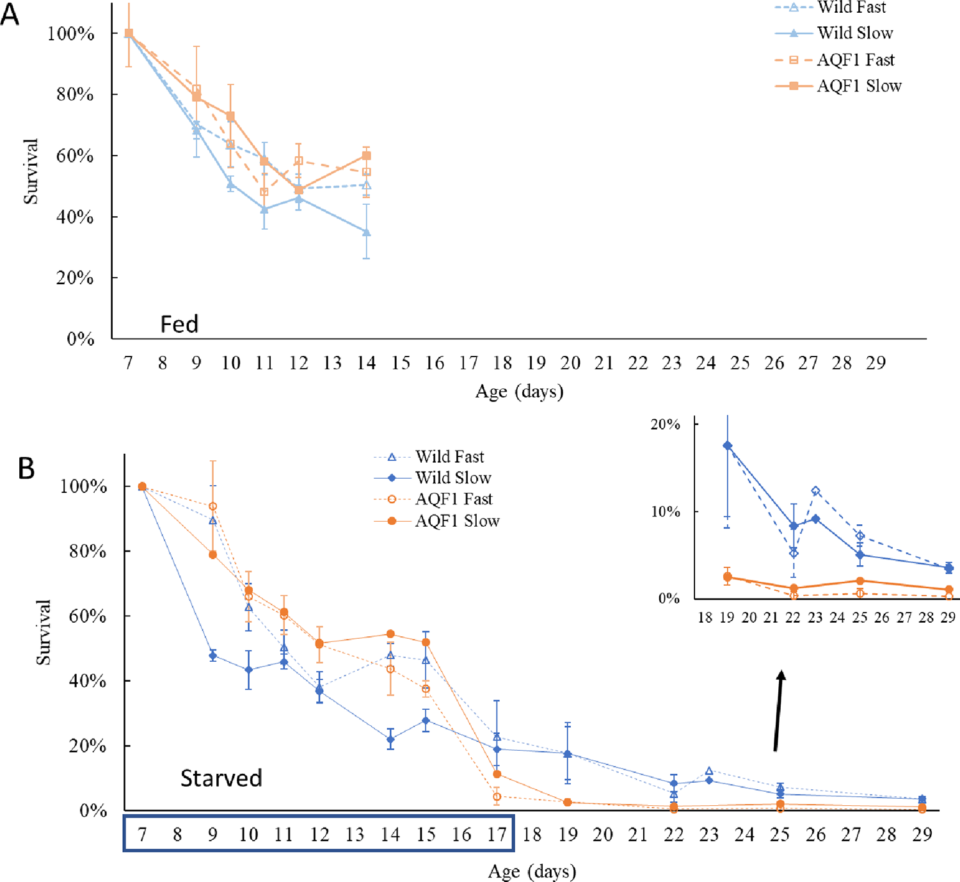
Larval survival and successful recruitment of subsequent generations following plantings of hatchery produced spat or adult oysters is an essential component to long-term restoration success. If planted oysters from selective breeding programs produce larvae that cannot survive the gauntlet of stressors in the estuarine environment, then long-term restoration success is stymied. While a 10-day period of no phytoplankton availability may be unlikely under natural conditions, patchiness in larval food quantity and nutritional quality are expected. Under these conditions, an ability to withstand and recover from periods with limited exogenous energy sources is a critical fitness trait.
Starvation tolerance is a complex trait, so we expected there might be interactions with growth rate. With the development of high size variance early in each line (typical of Eastern oysters), we separated each line into fast and slow early-growth cohorts to test for relationships with starvation tolerance.
When comparing starvation tolerance in larvae produced from wild parents and larvae produced from selectively bred parents, the selectively bred AQF1 line experienced nearly complete mortality during the last three days of starvation, while both fast and slow wild cohorts experienced better survival. Analysis of dissolved organic carbon suggests that although particulate food (phytoplankton) was withheld, micronutrients (e.g. dissolved organic matter) were present during starvation.

For the wild larvae in the fed treatment, the initial size separation between fast and slow growth cohorts was maintained throughout the experimental period, with no significant difference in growth rate observed. In the fed AQF1 line, the initial size distinction was maintained up to age 10 days (three days after size separation), but depressed growth of AQF1/Fast led to size convergence with AQF1/Slow by age 14 days. The reasons for changes in growth rate in AQF1/Fast (seen in both fed controls and the starved treatment) are not known, but they were apparently specific to a particular developmental stage and do not compromise the main finding of line growth rate parity.
The lower tolerance to starvation in the selected strain studied here is consistent with this trait being a costly adaptation weakened as a result of selection for fast growth or, more likely, a trait correlated with commercial traits under direct selection. Further comparisons with other Eastern oyster selected lines are needed to determine if this larval tolerance difference is a general result of domestication or specific to the tested lines.
Perspectives
For oysters, lines artificially selected for aquaculture support the oyster aquaculture industry, where lifetime fitness is not an objective. Rather, the emphasis is on farm-to-table production rates in which yield is largely a function of growth rate and survivorship. However, in the context of using hatcheries for stock enhancement, planted oysters must not only grow to maturity, but also successfully reproduce and generate offspring with robust abilities to withstand the many stressors encountered during the pelagic life stage.
At this early stage of selective breeding in Eastern oysters, the unknown phenotypic impacts from domestication selection may be large or small, depending on the trait. In our study, we showed a slightly lower tolerance to prolonged starvation in the AQF1 line intentionally selected for disease resistance over multiple generations of hatchery propagation. Replication of this experiment will be necessary using more biological replicates, different aquaculture lines and additional wild oysters to strengthen the inference that reduced larval starvation tolerance is linked to domestication selection generally.
Based on our initial results, we suspect bioenergetic processes related to micronutrient uptake and utilization may be promising candidate traits for investigating mechanistic changes as a result of domestication selection or are genetically correlated with other traits under direct selection.
The great successes of selective breeding to produce shellfish with improved aquaculture yields is likely to result in an increased reliance on selectively bred lines for oyster farming. With further oyster domestication expected, our work is of importance to understand inadvertent trait evolution as well as the potential impacts domesticated oysters can have on natural populations.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Authors
-

Corresponding author
Department of Natural Resources
Cornell University
Ithaca, New York, USA; and
Center for Environmental Science
University of Maryland
Cambridge, Maryland USA; and
NOAA Fisheries NEFSC
Milford Laboratory
Milford, Connecticut, USA -
Center for Environmental Science
University of Maryland
Cambridge, Maryland, USA -
Center for Environmental Science
University of Maryland
Cambridge, Maryland, USA; and
College of Earth, Ocean and Atmospheric Sciences
Oregon State University
Corvallis, Oregon, USA -
Department of Natural Resources
Cornell University
Ithaca, New York, USA
Related Posts

Health & Welfare
Avoiding genetic bottlenecks in broodstock selection
Broodstock selection programs for Atlantic halibut identify top first-generation performers to develop a strain best suited to aquaculture conditions.

Health & Welfare
Black tiger domestication, selective breeding advance in Australia
Using clear-water tank systems, CSIRO and a collaborating farm have advanced the domestication of black tiger stocks in Australia.

Health & Welfare
Genetic improvement of marine fish
Aquaculture often fails to enhance productivity and competitiveness with genetic improvement, perhaps due to uncertainty over which methods should be used.

Innovation & Investment
Unleashing the genetic potential of black tiger shrimp
The black tiger shrimp is the second most important, farmed shrimp species globally. Because of its importance in Australia, the new ARC Hub for Advanced Prawn Breeding will develop a scalable, advanced breeding program for the species as well as bring its genetic knowledge to a level comparable to that of livestock.