Understanding shrimp hemocytes
An important mechanism of defense against viruses, bacteria and other pathogens
Shrimp producers seeking to improve the profitability of their crops must be aware of external factors that may affect the survival and growth of their shrimp. One of the limiting factors for success in shrimp farming is the control of diseases. This control is mainly based on biosecurity, good nutrition and reduction of stress conditions during cultivation.
In this sense, the study of the immune system of marine shrimp stands out as a source of knowledge to determine the degree of the animal’s susceptibility and resistance to pathogenic microorganisms and parasites. In addition, these studies provide valuable knowledge to the relationship between the physical-chemical parameters of water and the degree of immune response.
The shrimp’s mechanisms of defense that allow the control of attacks by external agents such as viruses and bacteria include the production of hemocytes, defense cells present in the blood of shrimp. This is the central theme of this article, adapted and summarized from the original publication in Revista Acuacultura – National Chamber of Aquaculture, No. 126, December 2018.
Hemocytes: Cells to fight pathogens
White shrimp, like other invertebrate animals, depend on their immune system to protect themselves from diseases in situations where some microbe or foreign particle invades their tissues. This immunological reaction is manifested through cellular mechanisms where hemocytes play a very important role.
The blood of shrimp – or hemolymph – has a cellular component, which is the hemocytes, and a liquid component constituted of plasma that contains different humoral factors (macromolecules of the circulatory system). The immune-cellular and humoral responses act in an integrated manner creating defense mechanisms such as the coagulation of hemolymph; melanization by the prophenoloxidase (ProPO) system – a major, innate defense system in invertebrates; use of lectins for the recognition of foreign agents; and antibacterial, antifungal and antiviral peptide systems that act with interference RNA and a pattern of recognition proteins. We can also add the production of reactive forms of oxygen, phagocytosis and encapsulation, the latter two mainly performed by hemocytes (Iwanaga & Lee, 2005).
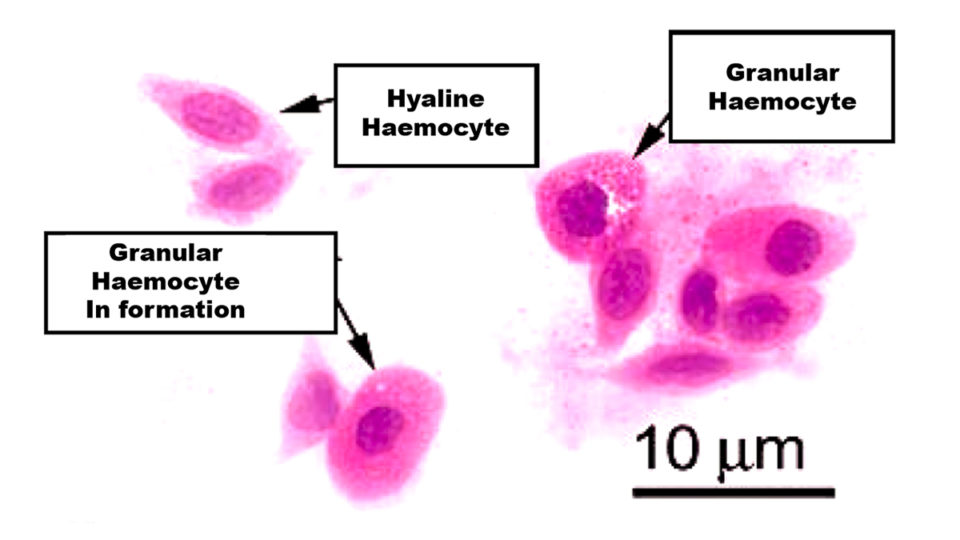
Hemocytes (Fig. 1) are produced in the hematopoietic tissues of shrimp, and there are two types:
- Hyaline hemocytes that absorb pathogens or foreign particles through the process of phagocytosis. They also intervene in the coagulation process.
- Granular hemocytes or granulocytes that – through encapsulation, nodule formation and cytotoxicity – destroy invading elements. They also intervene in melanization (ProPO system).
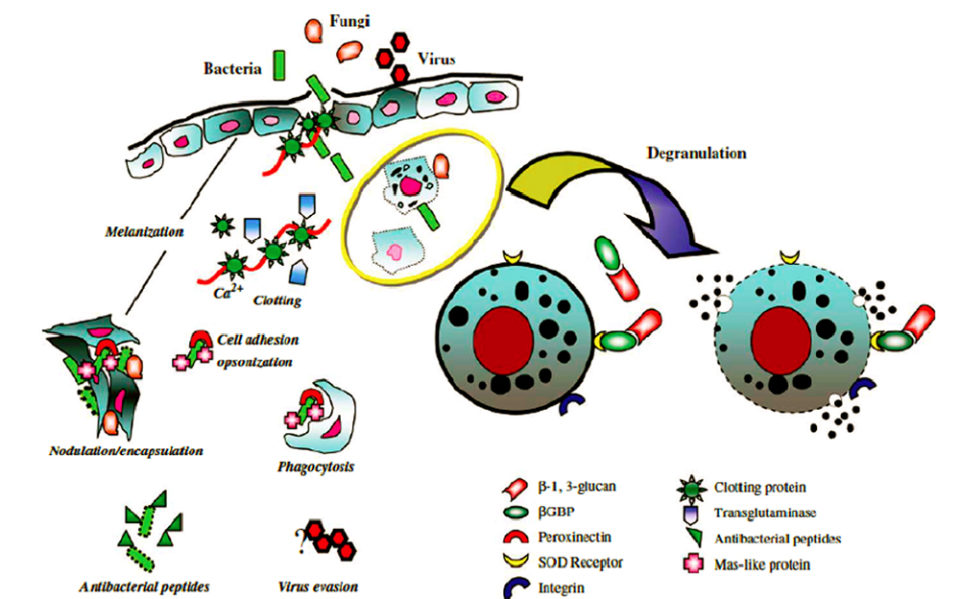
The hyaline hemocytes initiate the defense in the face of an injury with the coagulation process, a critical mechanism that protects the shrimp from an excessive loss of liquids as well as to capture and immobilize invading microbes. Next, the granular hemocytes secrete defensive enzymes that kill the microbes before being eliminated by other granulocytes through the processes of phagocytosis and/or encapsulation. Once the microbes are encapsulated or phagocytosed, the process of melanization, which is also led by the granular hemocytes, leaves them inert and prepares them to be expelled by the cuticular excretion or during the next molting cycle.
The hemocyte defense mechanism also causes the increase of cells with free radicals and stimulate the hyaline hemocytes to become granular hemocytes, increasing in this way the rate of elimination of pathogens by the degranulation process (Fig. 2).
Water quality and its effect on hemocyte production
The production of hemocytes as a response to disease attacks is closely linked to the physical-chemical parameters of water, such as temperature, oxygen, pH and salinity, as we will see below.
Relationship temperature – hemocytes during a white spot virus attack
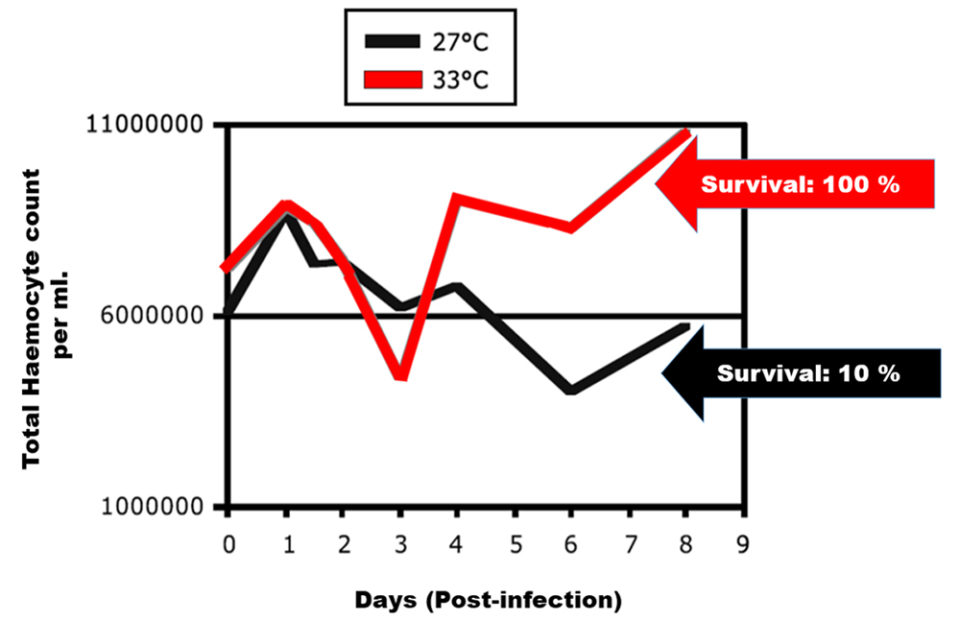
Experiments on the effect of temperature on shrimp infected with the white spot virus (WSSV) have shown that higher production of hemocytes occurs at higher temperatures (Sonnenholzner et al., 2002). Animals infected with WSSV achieved 100 percent survival when cultured at 33 degrees-C, while those cultured at 27 degrees-C only achieved 10 percent survival. This was due to the fact that, at 33 degrees-C, the production of hemocytes is greater (Fig. 3).
Another study that must be considered is the one by Wongmanneeprateep et al. (2010), which maintained juvenile shrimp infected with white spot at a constant temperature of 32 ± 1 degrees-C for seven consecutive days, thus eliminating WSSV infection. This study served as the basis for testing raceways where larvae infected with WSSV were cleaned after seven days at temperatures of 32 ± 1 degrees-C (Limsuwan, 2015, Fig. 4).
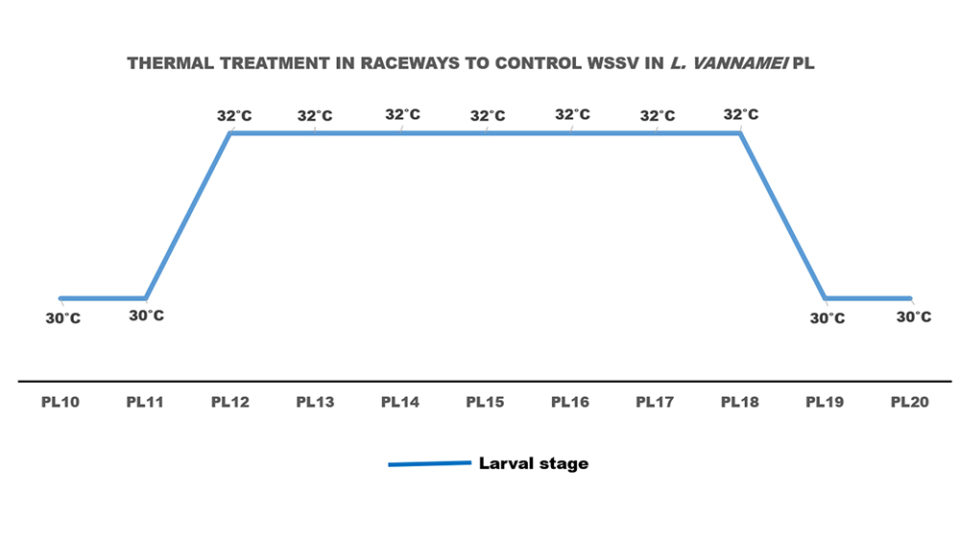
On the other hand, it must be considered that there is a risk in elevating the temperature above 30 degrees-C, as the populations of Vibrio bacteria can be increased (Chen et al., 2005), in which case the larvae can be dosed with shrimp health diets such as Nicovita Origin (composed of organic acids that produce a bactericidal effect on vibrios) and water bioremediation to control bacterial populations.
Concentration of dissolved oxygen and hemocyte production during a Vibrio spp. attack
The concentration of dissolved oxygen in water is one of the most important parameters in shrimp farming and is affected by environmental factors such as large plankton production and excessive accumulation of organic matter on pond bottoms. This increases the bacterial activity and leads to excessive oxygen consumption. Low levels of oxygen can affect the survival and growth of shrimp, because the reduced respiration and osmotic pressure of the shrimp at extremes can cause mortalities.
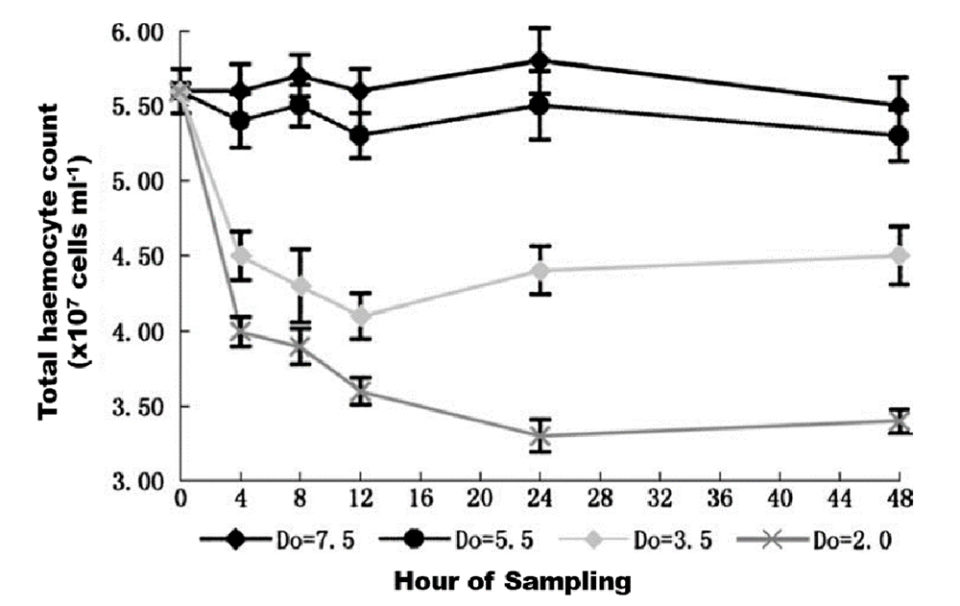
The production of hemocytes in marine shrimp depends on the concentration of oxygen in the water when pathogenic bacteria such as Vibrio spp. attack This is demonstrated by experiments carried out at different oxygen concentrations (7.5, 5.5, 3.5 and 2.0 ppm of dissolved oxygen in the water) (Fig. 5). The pathogenic bacteria Vibrio spp. could be controlled when the concentration of dissolved oxygen was at 5.4 and 7.5 mg/L, while the amount of hemocytes was relatively low at concentrations of 2.0 and 3.5 mg/L causing the shrimp to have very low defenses against these bacterial attacks (Ling-Xu Jiang et al., 2005).
Shrimp immune response at different pH levels
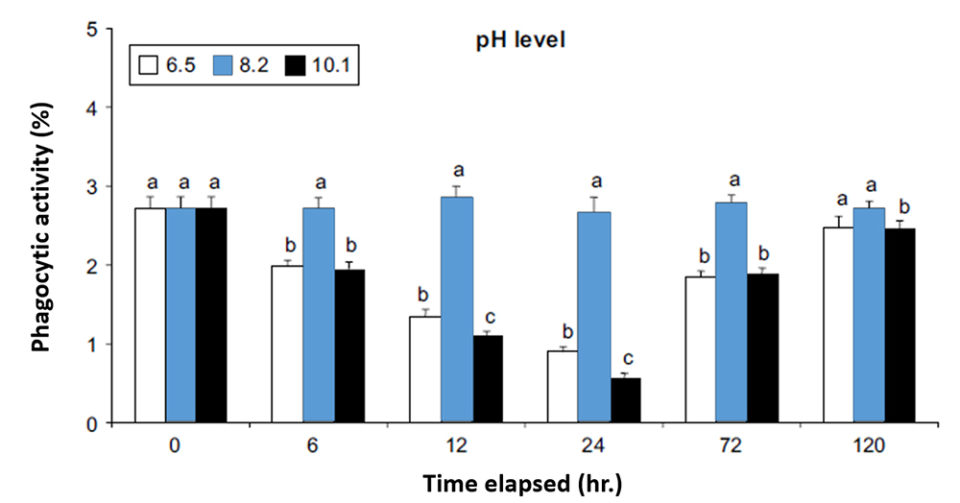
Studies conducted with L. vannamei shrimp exposed to a pathogenic strain of Vibrio alginolyticus (8.0 x 105CFU) at different pH levels showed differences in final survival (Li and Chen 2008). The lowest survivals were obtained at the lowest (6.5) and highest (10.1) pH, while the highest survival was obtained at pH 8.2. However, the pathogenic bacteria continued to reduce survival in all pH scenarios as the hours of exposure progressed (Fig. 6).

This study of the effect of pH on shrimp survival (Li and Chen, 2008) showed that, in shrimp culture, the most important parameter to stop vibriosis is the concentration of dissolved oxygen. However, if sudden changes in pH are experienced, shrimp survival could be reduced by reducing defense mechanisms such as phagocytic activity (Fig. 6), even with high concentrations of dissolved oxygen.
Sudden salinity changes decrease shrimp immune response
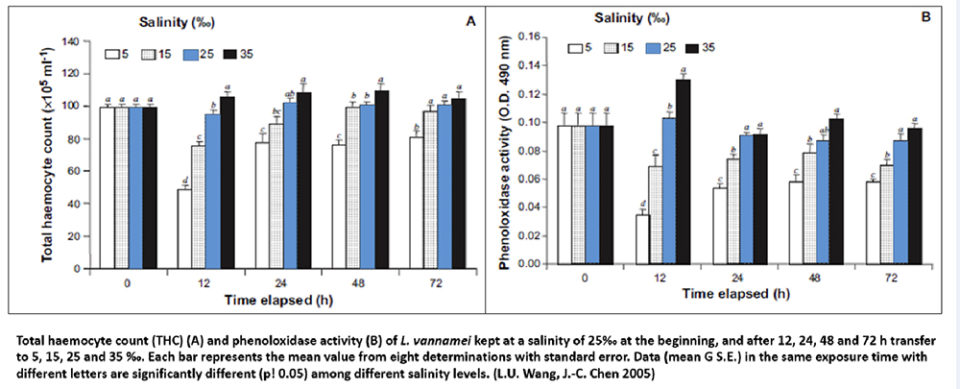
Experiments carried out with shrimp at 25 ppt salinity, injected with Vibrio alginolyticus (1.0 x 104CFU) and subsequently transferred to salinities of 5, 15, 25 (control) and 35 ppt for 24 to 96 hours, showed that the highest shrimp mortality occurred in those animals transferred from 25 to 5 ppt ‰ than for those shrimp transferred from 25 to 35 ppt. The shrimp were analyzed for total hemocyte count, phenol oxidase activity, respiratory burst, superoxide dismutase activity, phagocytic activity and bactericidal effect for V. alginolyticus.
Results indicate that when an abrupt change is made from a higher salinity (25 ppt) to a lower salinity (15 or 5 ppt), the immune defenses of the shrimp and their resistance to vibrios are diminished. Whereas if the shrimp in low salinity (25 ppt) are transferred to a high salinity (35 ppt), animal survival is not affected (Fig. 7). Therefore, an acclimation protocol is recommended that allows animals to have an appropriate time to osmoregulate with the water salinity without affecting their immune response (in the presence of vibrios), and, consequently, their survival.
Health diets complement the antibacterial action of hemocytes
In parallel to properly managing the aquatic environment during disease attacks for a greater production of hemocytes, it is advisable to complement these actions with the use of health diets such as Nicovita TERAP, whose antibacterial action is based on the use of organic acids that can inhibit and/or eliminate pathogenic bacteria in shrimp. In addition, the antibacterial action of these diets is also based on the action of its included immunostimulants, such as beta-glucans, nucleotides, selenium and zinc that increase the production of hemocytes in shrimp.
Thus, acting together through an improved aquatic environment and health diets as mentioned above, it is possible to achieve an effective control of pathogens such as bacteria Vibrio spp. which cause vibriosis, one of the most common and deadly diseases in cultured marine shrimp.
References available from author.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Author
-

Aquaculture Manager
Vitapro
Guayaquil, Ecuador
Related Posts
Health & Welfare
Emerging disease: Shrimp Hemocyte Iridescent Virus (SHIV)
SHIV is a new Pacific white shrimp virus in the Iridoviridae family. Authors also developed an ISH assay and a nested PCR method for its specific detection.

Health & Welfare
Natural strategies to improve growth and health of farmed shrimp
The application of probiotics, prebiotics and lipopolysaccharides has shown positive results with farmed shrimp, and is a very promising field, but more research is needed to clarify and foresee the molecular impact of these biological entities.
Health & Welfare
Study: TSV exposure may lessen YHV effects in white shrimp
Since Taura syndrome virus was in the Americas when yellow head virus become pandemic in Asia, it is possible TSV prevented shrimp in the Western Hemisphere from infection by materials from Asia containing YHV. To test this idea, the authors evaluated the viral interactions in shrimp preinfected with TSV and then challenged with YHV.

Health & Welfare
Aquaculture disease diagnosis is detective work
Those charged with determining the underlying cause of a disease outbreak have a wide variety of detective tools at their disposal.