Effect of full-fat, black soldier fly meal in juvenile Siberian sturgeon diets
Study evaluates physiological responses in first reported feeding trial using insect-based diets under aquaponics conditions

The Siberian sturgeon (Acipenser baerii) is one of the most valuable species in aquaculture, due to its production of caviar and high-quality meat for human consumption. Restocking programs are also of great interest for this species since it is included in lists of endangered wild populations. Compared to other sturgeon species, A. baerii shows rapid growth rate, resistance to pathogens, a relatively short reproductive cycle (seven to eight years) and can be reared using a wide range of diets and environmental conditions. Because of these features and a high commercial value, intensive aquaculture of Siberian sturgeons is presently carried out in different parts of the world and, in recent years, several studies regarding farming-related biology and nutritional requirements have been carried out.
Over the last years, several ingredients have been evaluated as fishmeal substitutes in sturgeon aquaculture diets, ranging from vegetable ones like soybean meal, rice concentrate, sesame oil cake, corn gluten and spirulina microalgae to animal ones such as poultry byproducts. Currently, with the goal of a further reduction of aquaculture’s environmental footprint, insect species like the black soldier fly (BSF; Hermetia illucens) represent very promising candidates as fishmeal alternatives. The significant interest in BSF meal as an aquafeed ingredient is due to their eco-friendly rearing in terms of land use, water consumption, carbon dioxide (CO2) emissions and on high feed conversion efficiency, as BSF larvae can grow on low-value organic byproducts, converting them into valuable biomass.
Furthermore, insects are part of the natural diet of Siberian sturgeon, and many insects have bioactive compounds like chitin, which at certain concentrations are able to boost the fish’s immune system and promote gut microbiota diversification. From the nutritional point of view, BSF larvae have a suitable protein content and the amino acid composition is similar to that of fishmeal. However, the BSF meal fatty acid profile has also some disadvantages, such as a high content of saturated fatty acids (SFA) and an extremely low content in polyunsaturated (PUFA) ones. PUFA are particularly important for fish since deficiencies in these compounds may cause a general deterioration of fish health, poor growth, low feed efficiency and often high mortality.
Some recent studies have tested different defatted BSF meal inclusion levels in aquafeed formulation for several fish species but results on fish physiological responses are still controversial. This topic, however, has scarcely been investigated in sturgeon aquaculture and most of the results are limited to zootechnical analyses. Nowadays, several laboratory approaches (histology, molecular biology, gas chromatography and infra-red spectroscopy) are available to evaluate fish welfare and quality and represent valid tools to assess the inclusion of new ingredients, like insect meal, in aquafeed production. In addition, the use of full-fat BSF meal is preferable to the highly defatted to reduce manufacturing costs.

This article – adapted and summarized from the original publication [Zarantoniello, M. et al. 2021. Physiological responses of Siberian sturgeon (Acipenser baerii) juveniles fed on full-fat insect-based diet in an aquaponic system. Sci Rep 11, 1057 (2021)] – reports on the results of a feeding study with Siberian sturgeon juveniles fed BSF diets in an aquaponic system.
Study setup
Our 60-day feeding trial was conducted at the aquaponics facility “Cooperativa Agricola Tanto Sole” (Treia, Macerata, Italy). Juvenile Siberian sturgeons, purchased from Azienda Agricola Pisani Dossi s.s., Cisliano, MI, Italy), were acclimated for one week in a single 500-liter tank equipped with mechanical, biological and UV filtration. Tank water temperature was 18 ± 0.5 degrees-C; ammonia and nitrite values were < 0.05 mg per liter, and nitrate was 10 mg per liter, all within acceptable sturgeon-rearing requirements. At the end of the acclimation period, fish were randomly allocated into six Media Based Aquaponic Systems (80 specimens per tank). Each aquaponic system consisted of a 1.56-square-meter hydroponic unit for plants cultivation and a 600-liter fish tank, for a total volume of 720 liters of water.
At the beginning of the experiment (t0), the six aquaponic systems were randomly assigned to the experimental groups (Hi0 or Hi50) according to an experimental design with triplicate tanks per dietary treatment. The feeding trial lasted for 60 days, during which the sturgeons almost triplicated their weight. Fish were fed as follows: fish fed on the 0 percent BSF meal diet (Hi0 group); and fish fed on the diet including 50 percent BSF full-fat prepupae [mature insect larvae] meal (Hi50 group). Feed particles were 0.5 to 1 mm in size. Sturgeons were fed the experimental diets (3 percent body weight daily) three times a day. At the beginning (t0) and at the end of the feeding trial (t1), after a 10-hour fasting period, the required fish were sampled, euthanized and properly stored for further analyses.
For detailed information on the experimental setup, design and fish husbandry; insects rearing and fish diet production; aquaponics system design and operation; data collection and histology; sturgeon microbiome, molecular and statistical analyses, refer to the original publication.
Results and discussion
The inclusion of BSF meal in aquafeed, as well as the physiological responses of fish, have been recently investigated in several important commercial species like Atlantic salmon (Salmo salar), rainbow trout (Oncorhinchus mykiss) and European sea bass (Dicentrarchus labrax).
But the results of our feeding study with Siberian sturgeon juveniles and enriched BSF meal – including zootechnical performances, fillet fatty acid composition, liver and gut integrity, expression of genes involved in fish growth, stress and immune response and gut microbiome – represent the first multidisciplinary investigation on the physiological effects of BSF-based diets in sturgeon juveniles. Furthermore, this is the first feeding trial using insect-based diets performed in an aquaponics system. This green technology combines aquaculture (production of fish) with horticulture (vegetables production), saving energy, water and nutrients, and representing an important step for sustainable aquaculture in a future zero-waste generation. Information available for the Siberian sturgeon and aquaponic systems is not available.
Previous studies highlighted that a defatted BSF prepupae meal dietary inclusion level higher than 25 percent impaired fish growth and welfare. Based on these results and previous studies which demonstrated the possibility to use higher full-fat dietary BSF meal inclusion levels after an enrichment procedure of the insect biomass, in our study we tested, for the first time, a 50 percent enriched full-fat BSF prepupae meal inclusion level in a practical diet for juvenile sturgeons in an aquaponic system; and we also applied a set of laboratory techniques to examine comprehensively the physiological responses of our experimental fish.
Our results showed that the inclusion of a 50 percent enriched full-fat BSF prepupae meal negatively affects fish growth and survival. These conclusions are supported by the expression of the growth markers analyzed and agree with previous studies reporting that inclusion of BSF meal levels higher than 40 percent often impairs fish growth and welfare. A possible explanation of these differences between the two experimental groups can be related to the fatty acid (FA) composition of the diets and the ability of sturgeons to biochemically convert some FA to others, and the related energy expenditure.
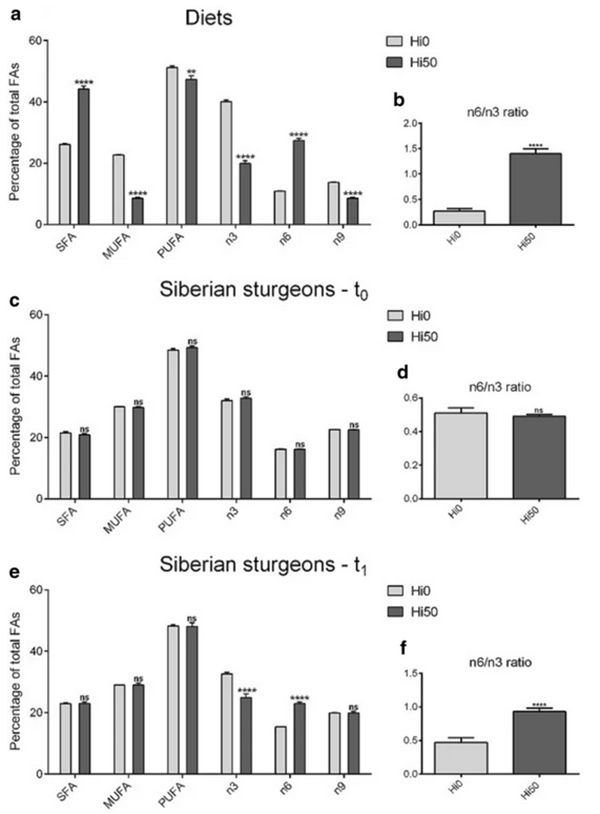
Also, we observed a lower diet acceptance was observed in the Hi50 group compared to Hi0, and the laboratory analysis performed suggested that the fish entered a fasting condition. Fasting is normally characterized by a growth reduction due mainly to a decrease in the production of certain genes involved in fish growth (IGFs). Accordingly, based on these and other observations, our results represent strong evidence that fish entered a fasting period. In addition, since malnutrition or fasting are nowadays considered stressors, this stressful situation is fully supported by the higher expression of a specific liver gene we detected in Hi50 compared to Hi0.
Histological analyses of intestinal tracts are also useful to provide evidence of fasting status in fish. Fasting is usually associated to changes in various intestinal membrane and cells, and our Hi50 group showed atrophy of intestinal mucosal folds and a dramatic decrease of specific types of intestinal cells compared to Hi0. Other researchers have reported no such negative effects after feeding trials with Siberian sturgeons fed diets with lower inclusion levels (37.5 percent) of defatted BSF meal.
Overall, our analyses of the intestine histology and expression of specific genes did not reveal any sign of inflammation in either of the experimental groups. Similar results were reported in other studies that showed a positive role of BSF meal dietary inclusion on fish gut welfare. BSF meal contains lauric acid and chitin, which have anti-inflammatory and immune-boosting properties. BSF meal is also known to rise biodiversity in the fish microbiome, necessary to improve fish health, metabolism, nutrition and immunity.
Perspectives
Our results demonstrated the feasibility of aquaponics systems for sturgeon culture and for testing new aquafeed ingredients like insect meal. However, the general low feed acceptance shown by the Hi50 group fish compared to the Hi0 group indicates the need of further studies aimed at improving the palatability of BSF-based diets intended for sturgeon culture.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Author
-

Corresponding author
Dipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, via Brecce Bianche, 60131, Ancona, Italy
Editor’s note: The original article has 17 co-authors.
Related Posts
Intelligence
An engineer’s design for a classroom aquaculture-aquaponics system
An aquaponics teaching system was designed, built and operated by students at the University of Arizona, integrating its operation and management into the educational curriculum. This engineering design will require minimum maintenance and will last years.

Responsibility
Haiti’s hatchery of hope
Built on a dream of feeding some of the world’s poorest and most vulnerable people, a charity-built tilapia hatchery in Haiti now learns to stand on its own.

Responsibility
A look at integrated multi-trophic aquaculture
In integrated multi-trophic aquaculture, farmers combine the cultivation of fed species such as finfish or shrimp with extractive seaweeds, aquatic plants and shellfish and other invertebrates that recapture organic and inorganic particulate nutrients for their growth.

Intelligence
City fish-farm prototype set to prove itself in Minnesota
The Urban Organics model of farming leafy greens in conjunction with fish can work in any city environment, the company says. The first step is destroying East St. Paul’s image as a food desert.