Can dietary inclusion of montmorillonite clay help mitigate the farmed shrimp AHPND pandemic?
Feeding trial results suggest the potential to improve growth, health

With the increasing intensification of culture systems, outbreaks of shrimp diseases are prevalent. In recent years, shrimp cultivation has suffered significant economic losses due to the emergence of a bacterial disease called acute hepatopancreatic necrosis disease (AHPND), which had caused and is still causing shrimp mortalities from 40 to 100 percent during the first 35–45 days after stocking shrimp postlarvae in grow-out ponds. An infectious disease spreading across countries and continents, AHPND is a bacterial pandemic in the shrimp farming industry.
Studies have revealed that Vibrio bacteria causing AHPND now show resistance to a wide range of antibiotics. With changing global regulatory trends on the use of antibiotics in animal farming, it is imperative to find alternatives to the prophylactic and therapeutic use of antibiotics in shrimp aquafeeds. Many feed additives with different modes of action directly or indirectly targeting the Vibrio bacteria and/or shrimp host are being suggested. An innovative idea in mitigating AHPND infection would be to directly bind the AHPND toxins, resulting in the toxins being removed from the body via fecal expulsion before they can migrate to the hepatopancreas to cause damage.
Clay minerals (e.g. montmorillonite) are commonly used as adsorbents to decrease the effects of dietary mycotoxins from animal feeds. A U.S. company (Amlan International Inc.), through its research collaborators, has discovered that its existing commercial calcium montmorillonite product (Calibrin®-Z, available in select international markets) can bind the AHPND PirAB toxins in vitro. We therefore decided to conduct a feeding trial to evaluate the effectiveness and efficacy of Calibrin-Z (CL) to mitigate AHPND.
Montmorillonite belongs to the smectite group of clay minerals and is essentially a hydrated calcium and/or sodium aluminosilicate. An important source of montmorillonite in nature is bentonite rock of clay. The main components of the montmorillonite used in this study were reported to be at least 70 percent montmorillonite and 15 percent amorphous hydrated silicon dioxide. It had been selected specifically for its ability to bind toxins and is processed with a proprietary heat treatment to activate broad-spectrum toxin binding. It has been shown to bind fungal and bacterial toxins in vitro and mitigate their effects in poultry and livestock.
![Ad for [BSP]](https://www.globalseafood.org/wp-content/uploads/2025/07/BSP_B2B_2025_1050x125.jpg)
We recently conducted a study to assess if, and to what extent, dietary CL inclusion could provide benefits for the growth performance, feed utilization and disease resistance of Pacific white shrimp (Penaeus vannamei) to AHPND. To the best of our knowledge, this is the first report of the effects of dietary montmorillonite on shrimp stomach microbiota during an AHPND infection. Considering the magnitude of the AHPND pandemic and the great interest from shrimp farmers in finding a cost-effective solution, investigating potential alternatives to antibiotics is pertinent and crucial.
Feeding trial and disease challenge
Four practical diets were produced using a commercial shrimp post-larval feed (Post Larva II crumbles #902, Gold Coin Specialities Ltd., Malaysia) as the base formulation. The commercial shrimp feed was ground into a fine powder before being re-pelleted. The positive (posCON) and negative (negCON) control diet consisted of the re-pelleted commercial shrimp feed with the addition of a pellet binder (Pegabind®, Bentoli AgriNutrition Co., Ltd., Thailand). Only the shrimp fed the posCON diet was later challenged with Vibrio parahaemolyticus (VPAHPND). Two other diets with the inclusion of 0.25 percent (0.25 percent CL) or 0.5 percent (0.5 percent CL) montmorillonite clay (Calibrin®-Z) at the expense of α-cellulose were also pelleted. The control diets did not have any added clay.

At the start of the feeding trial, all SPF shrimp (PL30) were starved overnight, and groups of 20 apparently healthy shrimp were weighed, transferred to each of the 15-liter aquarium and randomly assigned to one of the dietary treatments. Triplicate groups of shrimp were hand-fed with their respective diet three times daily at 9 a.m., 2 p.m. and 6 p.m. to apparent satiation for two weeks. All shrimp were counted and batch-weighed at the end of the feeding trial. Throughout the feeding trial, the salinity, dissolved oxygen, pH and temperature of culture water were within the range of 23 ± 1‰, 5.5 ± 0.1 mg/L, 8.1 ± 0.2, 28 ± 0.4 degrees-C, respectively.
After the growth trial was completed, a sub-sample of the required number of shrimp from each dietary treatment were randomly removed from each aquarium, pooled and then re-stocked at 16 shrimp per aquarium for the disease challenge trial. The disease challenge was carried out using the immersion method. Shrimp from each aquarium in the posCON, 2.5 percent CL or 5 percent CL groups were placed for 15 minutes (with aeration) in a 1-liter Erlenmeyer flask containing 150 mL of VPAHPND culture (108 CFU/mL). Then, the bacterial suspension including shrimp was poured directly back into their respective aquarium containing clean artificial seawater to obtain a bacterial density of about 106 CFU/mL. The negative control treatment (negCON) was included to serve as an environmental control and the shrimp were not challenged with VPAHPND but immersed in sterile tryptic soy broth plus 1.5 percent NaCl. During the seven-day disease challenge trial, shrimp were fed two times daily their respective diet and received a 50 percent water exchange in the morning. All shrimp were observed every three hours to monitor gross clinical signs and mortality. Dead shrimp were recorded and removed from the experimental tanks.
After the AHPND challenge, the hepatopancreas was aseptically excised for microbiological and histopathological analysis. The shrimp stomach was also removed for microbiota analysis. All data collected were then subjected to statistical analysis. Comprehensive details on the methodologies used can be found in the original publication by Ng et al. (2023), Dietary montmorillonite clay improved Penaeus vannamei survival from acute hepatopancreatic necrosis disease and modulated stomach microbiota. Animal Feed Science and Technology 297, 115581.
The efficacy of montmorillonite clay (Calibrin®-Z)
All experimental diets were well received by the shrimp, and by the end of the feeding trial, shrimp more than doubled their initial body weight. The inclusion of dietary CL did not negatively influence growth, feed utilization efficiency, feed intake and survival of shrimp, and all parameters were not significantly different (P > 0.05). Feed conversion ratio (FCR) was excellent with values close to 1.0 and shrimp survival was 95 percent or higher in all treatment groups.
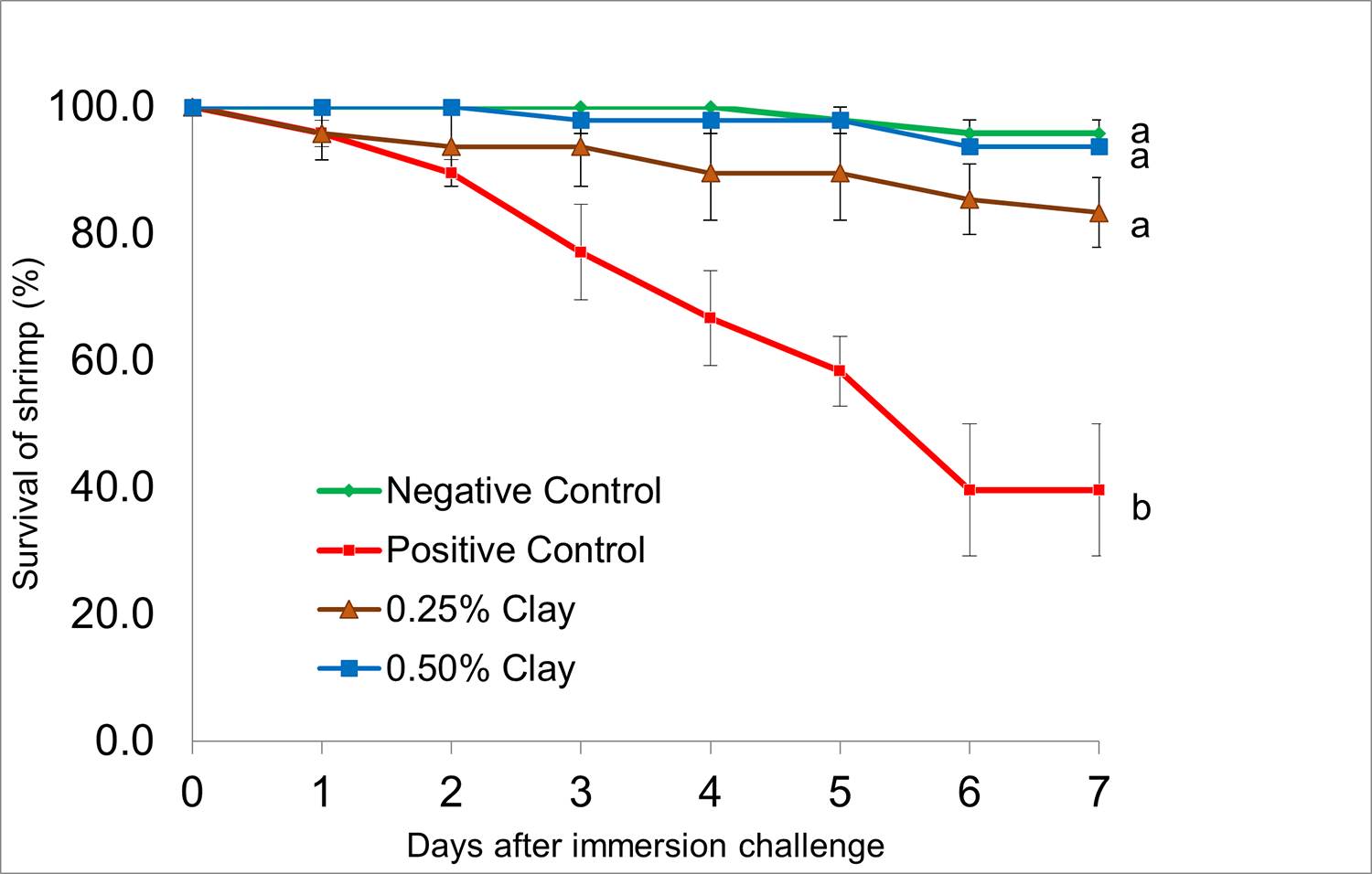
Shrimp in the negCON group (not challenged with VPAHPND) showed an average survival of 95.8 ± 2.1 percent over the seven days of the disease challenge trial. There were only two shrimp mortalities out of a total of 48 shrimp in the negCON group. Shrimp in the VPAHPND-challenged posCON group had the lowest average survival of 39.6 ± 10.4 percent, which was significantly lower (P < 0.05) compared to the negative control. The infected shrimp showed typical clinical signs of AHPND. The survival of shrimp fed CL-supplemented diets, irrespective of dietary level, were not significantly different compared to the unchallenged negCON group. There were no significant differences in survival of shrimp fed either 0.25 percent or 0.5 percent CL during the disease challenge trial.
Stomach microbiota
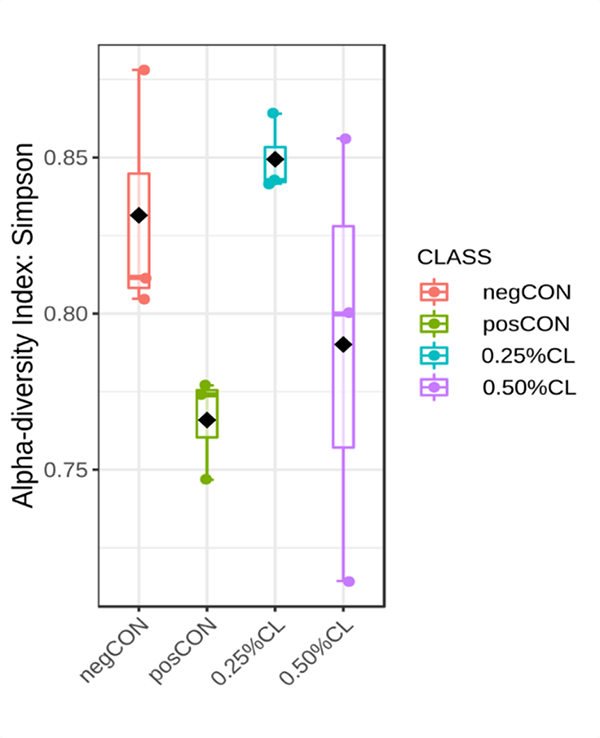
The Simpson index of the alpha-diversity plot measures the richness (number of amplicon sequence variant, ASV) and the evenness of ASV making up the richness of the sample. A high Simpson index indicates high number of ASV and more similar proportions of ASV. When challenged with VPAHPND, it was observed that the bacterial diversity in the posCON group was depressed compared to the unchallenged healthy shrimp in the negCON group. Supplementing shrimp diets with 0.25 percent CL tended to restore stomach bacterial richness and diversity index much more than the 0.5 percent CL group.
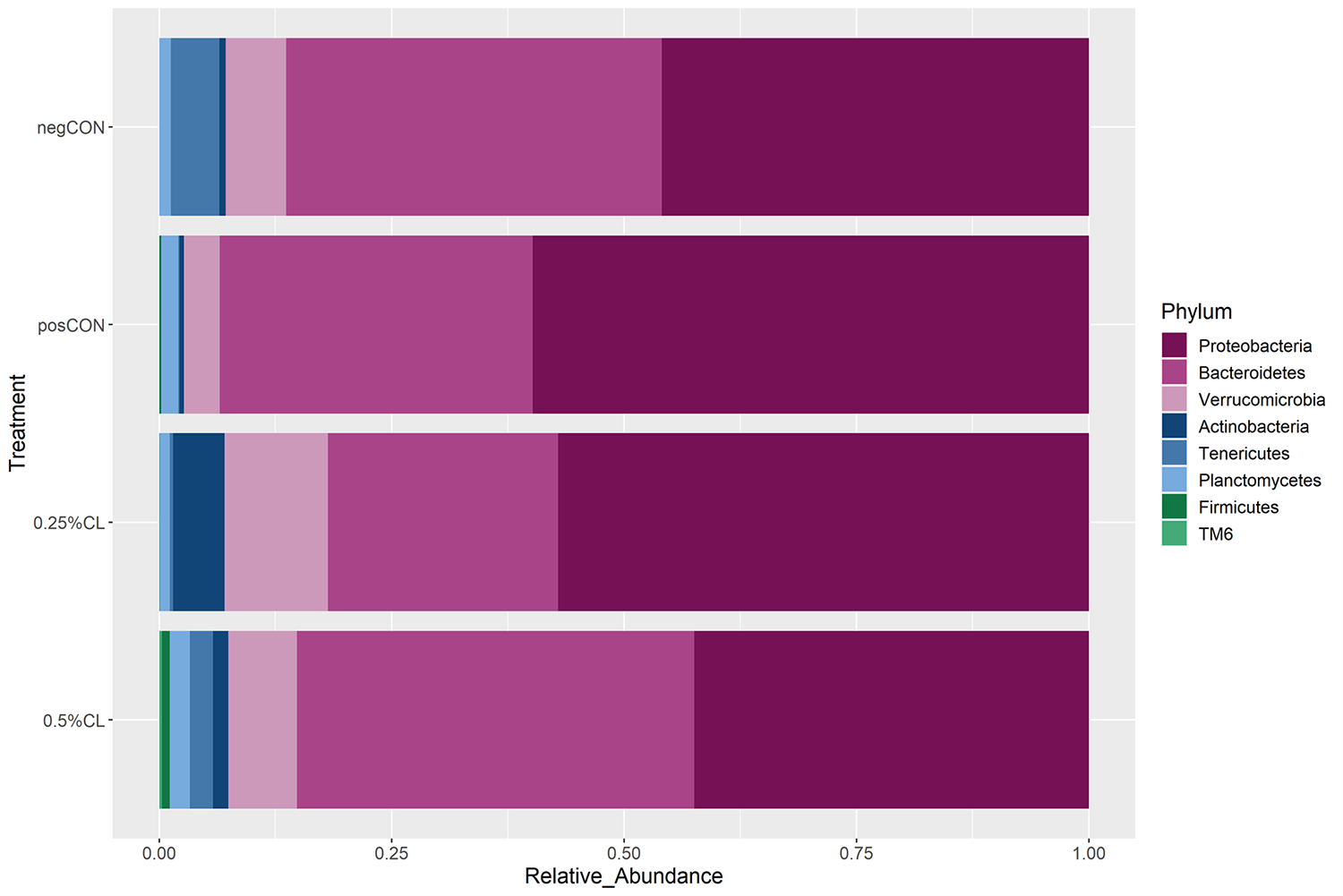
At the phylum level, shrimp stomach bacterial community structure consisted mainly of Proteobacteria and Bacteroidetes with Verrucomicrobia a distant third in relative abundance. The accumulated abundance of these three phyla was higher than 90 percent across treatment groups. The posCON group had increased relative abundance of Proteobacteria and decreased relative abundance of Tenericutes compared to the negCON group. Increased relative abundance of Verrucomicrobia and Actinobacteria was observed in the stomach of shrimp fed the CL-supplemented diets. At the 0.5 percent CL treatment group, slightly more abundance of Firmicutes was observed.
The potential of montmorillonite clays
The use of montmorillonite as an ingredient in livestock feeds has generally reported beneficial results. Very little information is currently available on the impact of dietary montmorillonite on growth performance of aquatic animals. Apart from species-specific differences, the discrepancies among various published research on the effects of montmorillonite on growth performance of aquatic animals are most likely due to the source and type of montmorillonite used. The purity, chemical composition, and functional properties of natural montmorillonite are very much dependent on the place where the clay mineral was mined. Calibrin®-Z is unique in its mining location and processing that it may not be possible to extrapolate its positive effects as observed in the present study to all montmorillonites.
When challenged with VPAHPND, survival of shrimp fed 0.25 percent or 0.5 percent CL was high (83 to 94 percent) and was not significantly different compared to the unchallenged negative control group. This result suggests that dietary montmorillonite was able to protect shrimp from mortality caused by bacterial toxins released by VPAHPND.
We believe that the main mode of action of montmorillonite in mitigating AHPND is through the adsorption of AHPND’s binary PirAB toxins on its surfaces and interlayers. Bacterial toxins can migrate through interconnected pores and capillary channels towards internal binding sites. Various positively charged sites in the montmorillonite structure such as the interlayer cations and edges of the octahedral units provide the adsorption sites. The naturally occurring opal lepispheres help maintain the layered sheet structures of CL and contribute to the high binding capacity.
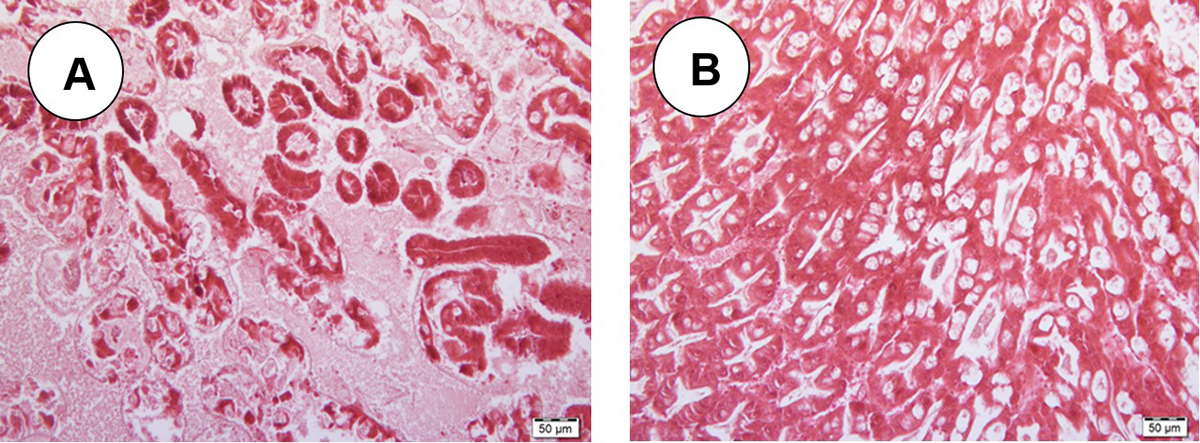
Adsorptive binding forces on clay surfaces include hydrophobic interactions, chelation, hydrogen bonding, electrostatic attractions and van der Waals forces. In the present study, we believed that the adsorbed bacterial toxins by montmorillonite were rendered unavailable for absorption in the shrimp gut and eventually expelled from the body through fecal excretion. Furthermore, histopathological examination of the hepatopancreas, the main target of the PirAB toxins, confirmed that shrimp fed montmorillonite -added diets showed very little damage compared to AHPND-challenged shrimp fed the control diet without any added montmorillonite. This showed that little or no PirAB toxins migrated to the hepatopancreas to cause tissue damage as the adsorbed toxins by MMT were likely moved along the gut away from the hepatopancreas by peristalsis and eventually excreted via fecal matter.
The present study is the first report on the modulation of shrimp stomach microbiota by dietary montmorillonite during a disease infection. The addition of montmorillonite in the diet of white shrimp was observed to increase microbiota diversity, especially in the 0.25 percent CL group. Stomach microbial diversity of infected shrimp was markedly reduced by AHPND compared to healthy shrimp. Not much information is currently available on stomach microbiota changes in response to AHPND, but in general, it is known that a higher gut bacterial diversity reflects better gut health and subsequent animal health.
At the phylum level, we identified Proteobacteria and Bacteroidetes as the main groups. We observed that under AHPND infection, the relative abundance of Proteobacteria was increased as compared to unchallenged shrimp. We identified bacteria of the genus Pseudoalteromonas, Tenacibaculum and Marinimicrobium as members of the healthy microbiome and Lysobacter as being relatively enriched in the stomach of AHPND-infected shrimp. The actual roles played by these bacterial community changes remain to be elucidated.
Shrimp fed 0.25 percent CL showed the highest relative abundance of Verrucomicrobia and Actinobacteria, followed by the 0.5 percent CL group. Firmicutes relative abundance was more prevalent in the 0.5 percent CL group compared to other treatment groups. A full description of the stomach microbiota results can be obtained from the original publication. Apart from the ability of montmorillonite to bind PirAB toxin and mitigate AHPND-inflicted stomach dysbiosis, the ability of dietary montmorillonite in modulating stomach microbiota might also be due to the ability of montmorillonite to adsorb and immobilize bacteria selectively on their surfaces and interlayers giving rise to different bacterial profiles observed in this study.
Perspectives
In conclusion, dietary montmorillonite can be used as an alternative to antibiotics for the mitigation and prevention of the AHPND pandemic in shrimp aquaculture. It provides a natural solution to a present critical problem in shrimp farming sustainability. In the present study, we intentionally used shrimp sizes that are known to be most vulnerable to AHPND in the production cycle. Based on the laboratory results, we would suggest that CL at dietary levels of at least 0.25 percent be added in commercial shrimp feeds as an “insurance policy” and preventative measure against production and economic losses from potential AHPND outbreaks in shrimp farms. Further longer-term and field-based studies are warranted to fully elucidate the beneficial impact of montmorillonite in the feeds of farmed aquatic animals.
References available from the author.
Now that you've reached the end of the article ...
… please consider supporting GSA’s mission to advance responsible seafood practices through education, advocacy and third-party assurances. The Advocate aims to document the evolution of responsible seafood practices and share the expansive knowledge of our vast network of contributors.
By becoming a Global Seafood Alliance member, you’re ensuring that all of the pre-competitive work we do through member benefits, resources and events can continue. Individual membership costs just $50 a year.
Not a GSA member? Join us.
Author
-

Corresponding author
Retired Professor of Aquaculture Nutrition, School of Biological Sciences, Universiti Sains Malaysia, Penang, Malaysia; and Asian Aquafeeds Services, Penang, Malaysia
Related Posts

Health & Welfare
Updates on shrimp diseases AHPND, NHP at Aquaculture America 2018
Several shrimp-disease papers were presented at Aquaculture America 2018, including one on the detection of AHPND at a site in the United States.

Health & Welfare
Challenging Pacific white postlarvae with AHPND
Study results indicate that P. vannamei challenged with AHPND in biofloc had higher survival rates than shrimp challenged in clear water.

Health & Welfare
Clinical case report: EMS/AHPND outbreak in Latin America
Behind the successful control of Early Mortality Syndrome (EMS), or Acute Hepatopancreatic Necrosis Disease (AHPND), in a Latin American shrimp nursery.

Health & Welfare
Quality, survival of L. vannamei offspring from ablated, non-ablated females
Evaluating the quality and survival of Pacific white shrimp postlarvae from non-ablated female broodstock, a holistic biosecurity and management strategy.